While glycolysis is highly conserved between unicellular yeast and multicellular humans, it appears that not only the glycolytic functions but also the secondary (aka “moonlighting”) functions of the relevant proteins remain largely consistent. This consistency presents unique opportunities for better understanding human glycolysis in muscle tissue and elsewhere.
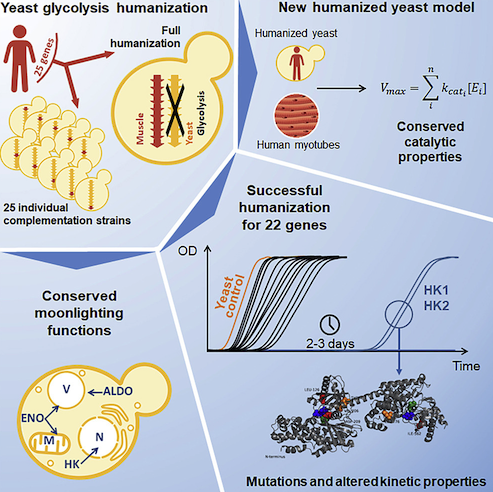
The technical challenges of studying glycolytic enzymes are not trivial—largely due to genetic redundancy for such critical components—but a recent report by Boonekamp et al. in Cell Reports explains how the authors built on previous work to overcome the obstacles in yeast. Employing a minimized set of glycolytic genes that were also relocated to a single chromosome (strain SwYG) made it possible to swap in human orthologs with relative ease.
The authors first looked at direct complementation of yeast genes by 25 human glycolytic enzymes. Remarkably, 22 of 25 human genes readily complemented their yeast counterpart. The three exceptions were hexokinases 1, 2, and 3 (HsHK1, HsHK2, and HsHK3), which are roughly twice the size of their yeast orthologs and have lower sequence conservation.
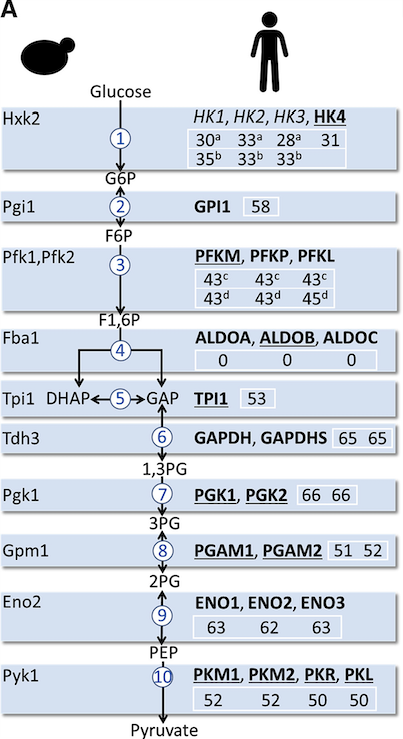
As glycolysis cannot take place without the hexokinases, the authors first looked more closely at HsHK1 and HsHK2 to ask why they fail to function in yeast. Upon exposure to glucose, for which the native human proteins could not support growth, they created conditions to select for systematic mutations that conferred improved glycolysis. The mutant proteins proved to be less sensitive to inhibition by glucose-6-phosphate (G6P), a potent allosteric inhibitor of hexokinase activity.
With this understanding, the authors set out to complement the entire pathway in yeast with human proteins. They created two different strains, one with HsHK2 because it is considered the main isoenzyme in human muscle (strain HsGly-HK2), and one with HsHK4 because it shows less inhibition by G6P (strain HsGly-HK4). The HsGly-HK2 strain could not grow well on glucose until after a long lag phase in which mutations were selected near the G6P-binding site of HsHK2.
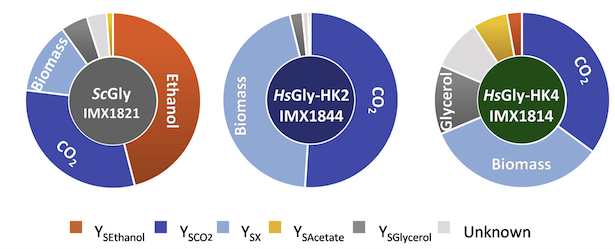
Comparing the uptake and output of the two humanized yeast strains to the native yeast strain revealed a number of intriguing differences, especially around the different enzyme activities between human and yeast. These differences led to different behaviors on different carbon sources and overall slower growth and glycolytic flux for the humanized strains versus native yeast.
Three glycolytic yeast enzymes have secondary “moonlighting” roles beyond their function in glycolysis. Hexokinases are involved in glucose repression of genes such as invertase (SUC2), and, indeed, it appears that human hexokinases can also at least partially complement this secondary function. Yeast aldolase (FBA1) plays a secondary role in vacuolar function that is required for growth at alkaline pH. This function is likewise complemented by the human orthologs, where the humanized strains can grow at pH 7.5. The third moonlighter is enolase, where yeast ENO2 is required for mitochondrial import of tRNALys, thereby allowing growth at higher temperatures and on non-fermentable carbon sources. All three human enolases can at least partially complement this growth defect, and thus appear to have the same secondary function.
Despite the high conservation of functions, the humanized yeast strains have a slow growth phenotype. The authors used this phenotype to employ adaptive laboratory evolution to see which genomic changes restore growth. Interestingly, the mutations that restored growth were mostly not in the glycolytic enzymes themselves, but in associated factors that regulate enzyme abundance and activity. The identity of these regulators and the potential for targeting them in human muscle have already rewarded the successful transfer of skeletal muscle glycolysis into yeast.