Cells are efficient machines, honed by selection. Recent studies are showing us how cells are more efficient than we realized, with proteins discovered to have multiple functions that can be surprisingly different under changes in environment. A recent paper in Molecular Cell describes Get3p as one such protein. Under non-stress conditions, Get3p acts as an ATP-dependent guide for post-translational insertion of tail-anchored proteins into membrane. Under stress, however, Get3p morphs into an antioxidant “guard” protein that prevents toxic protein aggregation by functioning as a chaperone. What are the mechanics of such a switch?
The finding came about in something of a backward way, where Ulrich et al. set out to find a eukaryotic chaperone protein specifically activated during oxidative stress. They looked for similarity to Hsp33, a bacterial protein that protects against oxidative damage by activation of a zinc-coordinating, four-cysteine motif. Their search led them to Get3p, which shares little overall homology but has the same conserved CXC and CXXC motifs.
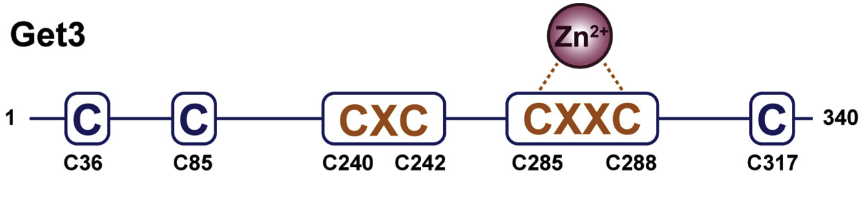
Whereas the bacterial protein acts only in stress, yeast Get3p also has a non-stress role that differs from its role in stress response. Under non-stress conditions, it is the CXXC motif that plays the major role, where the two cysteines of the motif bind zinc, which aids the protein in dimerization. In the switch to the chaperone-active form, however, it is the nearby CXC motif that plays the critical role.
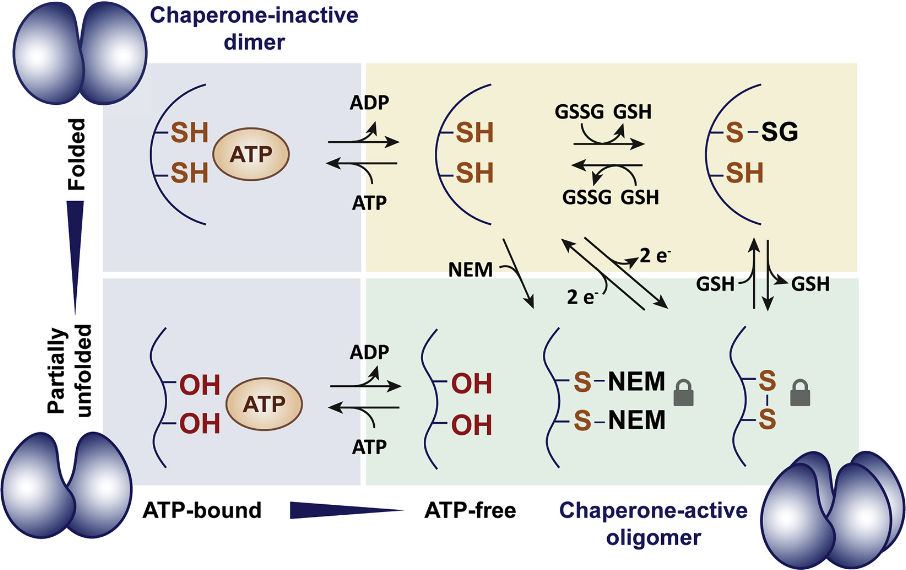
During oxidative stress, the switch to the chaperone form of the Get3p protein requires two things: (1) loss of bound ATP (with accompanying loss of ATPase activity, see below); and (2) oxidation of the free thiol groups (-SH) on residues C240 and C242 of the CXC motif. Once oxidized from thiol groups to sulfur groups, the two sulfurs of the CXC can form an intramolecular disulfide bond. Thus, in the presence of an ROS like H2O2, the new bond reconfigures the protein to become a non-ATP-requiring active chaperone. The CXC motif, then, acts as a “redox” switch.
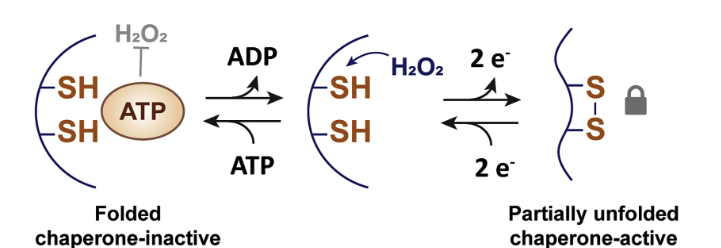
The role of ATP with respect to the redox switch is illustrated in the above schematic, where the presence of ATP makes the two thiol groups inaccessible to oxidation. When ATP is lost (due to the oxidative stress that reduces cellular ATP levels), the -SH groups can be oxidized to -S groups that bind to one another and partially unfold the protein, making it chaperone active.
The authors further show how the CXC motif is dispensable for the normal protein-targeting function of Get3p but absolutely required for the chaperone function. Thus, Get3p has two cysteine motifs that act in fully different ways, where CXXC is required for normal guide function while CXC is required for protection from oxidative stress.
Whereas the toxicity of reactive oxygen species is well established, the elucidation of a redox switch-triggered stress response in yeast provides an important clue to how eukaryotes detoxify these oxidants. It would not be surprising to find other cysteine-motif proteins acting as antioxidants in related processes that save cells from damage. As usual, yeast is an excellent model for understanding these intricate relationships.